Цикл кори молочная кислота
В клетках организма всегда существует потребность в глюкозе:
- для эритроцитов глюкоза является единственным источником энергии,
- нервная ткань потребляет около 120 г глюкозы в сутки и эта величина практически не зависит от интенсивности ее работы. Только в экстремальных ситуациях (длительное голодание) она способна получать энергию из неуглеводных источников (кетоновые тела),
- глюкоза играет весомую роль для поддержания необходимых концентраций метаболитов цикла трикарбоновых кислот (в первую очередь оксалоацетата).
Таким образом, при определенных ситуациях – при низком содержании углеводов в пище, голодании, длительной физической работе, т.е. когда глюкоза крови расходуется и наступает гипогликемия, организм должен иметь возможность синтезировать глюкозу и нормализовать ее концентрацию в крови. Это достигается реакциями глюконеогенеза.
По определению, глюконеогенез – это синтез глюкозы из неуглеводных компонентов: лактата, пирувата, глицерола, кетокислот цикла Кребса и других кетокислот, из аминокислот.
Необходимость глюконеогенеза и его значение для организма демонстрируют два цикла – глюкозо-лактатный и глюкозо-аланиновый.
Глюкозо-лактатный цикл (цикл Кори)
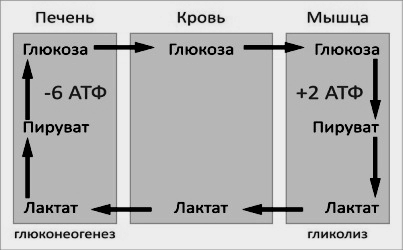
Глюкозо-лактатный цикл – это циклический процесс, объединяющий реакции глюконеогенеза и реакции анаэробного гликолиза. Глюконеогенез происходит в печени, субстратом для синтеза глюкозы является лактат, поступающий в основном из эритроцитов или мышечной ткани.
В эритроцитах молочная кислота образуется непрерывно, так как для них анаэробный гликолиз является единственным способом образования энергии.
В скелетных мышцах высокое накопление молочной кислоты (лактата) является следствием гликолиза при очень интенсивной, субмаксимальной мощности, работе, при этом внутриклеточный рН снижается до 6,3-6,5. Но даже при работе низкой и средней интенсивности в скелетной мышце всегда образуется некоторое количество лактата.
Убрать молочную кислоту можно только одним способом – превратить ее в пировиноградную кислоту. Однако сама мышечная клетка ни при работе, ни во время отдыха не способна превратить лактат в пируват из-за особенностей изофермента лактатдегидрогеназы-5. Зато клеточная мембрана высоко проницаема для лактата и он движется по градиенту концентрации наружу. Поэтому во время и после нагрузки (при восстановлении) лактат легко удаляется из мышцы. Это происходит довольно быстро, всего через 0,5-1,5 часа в мышце лактата уже нет. Малая часть молочной кислоты выводится с мочой.
Большая часть лактата крови захватывается гепатоцитами, окисляется в пировиноградную кислоту и вступает на путь глюконеогенеза. Глюкоза, образованная в печени, используется самим гепатоцитом или возвращается обратно в мышцы, восстанавливая во время отдыха запасы гликогена. Также она может распределиться по другим органам.
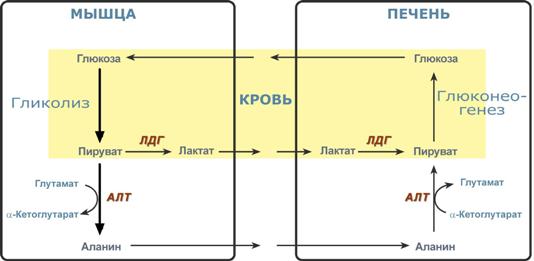
Глюкозо-лактатный (выделен желтым) и глюкозо-аланиновый циклы
Глюкозо-аланиновый цикл
Целью глюкозо-аланинового цикла также является уборка пирувата, но кроме этого решается еще одна немаловажная задача – доставкааминного азота из мышцы в печень.
При мышечной работе и в покое в миоците распадаются белки и образуемые аминокислоты трансаминируются с α-кетоглутаратом и полученный глутамат взаимодействует с пируватом. Образующийся аланин является транспортной формой аминного (аминокислотного) азота и пирувата из мышцы в печень. В гепатоците идет обратная реакция трансаминирования, аминогруппа через глутамат передается на синтез мочевины, пируват используется для синтеза глюкозы.
Кроме мышечной работы, глюкозо-аланиновый цикл активируется во время голодания, когда белки мышц и других тканей распадаются и многие аминокислоты используются в качестве источника энергии, а их азот необходимо доставить в печень.
Источник
Цикл
Кори (глюкозо-лактатный цикл) открыла
чешская ученая, лауреат Нобелевской
премии Тереза
Кори.
Он представляет собой биохимический
транспорт лактата
из мышц в печень и дальнейший синтез
глюкозы
из лактата, катализируемый ферментами
глюконеогенеза:
При
интенсивной мышечной работе и в условиях
отсутствия или недостаточного числа
митохондрий
(например, в эритроцитах)
глюкоза
подвергается анаэробному гликолизу
с образованием лактата.
При накоплении лактата в мышцах возникает
лактат-ацидоз, раздражаются чувствительные
нервные окончания, что вызывает боль в
мышцах.
Лактат
переносится кровью в печень и превращается
в пируват, а затем в глюкозу (глюконеогенез),
которая с током крови может возвращаться
в работающую мышцу.
Направление
лактатдегидрогеназной реакции в
работающих мышцах и печени обусловлено
отношением концентраций восстановленной
и окисленной форм НАД+:
отношение НАД+/НАДН
в сокращающейся мышце больше, чем в
печени.
7.7. Спиртовое брожение
Спиртовое брожение
осуществляется дрожжеподобными
организмами, а также некоторыми плесневыми
грибками:
![]()
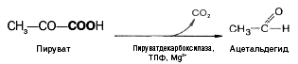
Механизм
реакции близок к гликолизу. Расхождение
начинается после этапа образования
пирувата. При гликолизе пируват при
участии фермента ЛДГ и кофермента НАДН
восстанавливается в лактат. При спиртовом
брожении пируват подвергается
декарбоксилированию, в результате
образуется ацетальдегид, а затем при
восстановлении его — этанол:

При молочнокислом
брожении ПВК не декарбоксилируется, а,
как и при гликолизе в животных тканях,
восстанавливается при участии ЛДГ за
счет водорода НАДН.
7.8. Пентозофосфатный путь превращения глюкозы
Пентозофосфатный
путь –
альтернативный путь окисления глюкозы.
Его функции:
—
поставляет клеткам кофермент НАДФН,
использующийся как донор водорода в
реакциях восстановления;
—
обеспечивает клетки пентозофосфатами
для синтеза нуклеотидов и нуклеиновых
кислот.
Пентозофосфатный
путь не приводит к синтезу АТФ.
Ферменты пути
локализованы в цитозоле.
В
пентозофосфатном пути превращения
глюкозы выделяют окислительный и
неокислительный пути образования
пентоз.
Окислительный
путь включает
две реакции дегидрирования. Коферментом
дегидрогеназ является НАДФ+,
восстанавливающийся в НАДФН. Пентозы
образуются при окислительном
декарбоксилировании.
Неокислительный
путь включает
реакции переноса 2- и 3-х углеродный
фрагментов с одной молекулы на другую.
Этот путь служит для синтеза пентоз.
Процесс обратим, и из пентоз могут
образовываться гексозы.
Пентозофосфатный
путь образования пентоз протекает в
печени, жировой ткани, молочной железе,
коре надпочечников, эритроцитах.
1).
Дегидрирование глюкозо-6-фосфата при
участии глюкозо-6-фос-фатдегидрогеназы
и кофермента НАДФ+
с образованием 6-фосфоглюконо--лактона
и НАДФН:
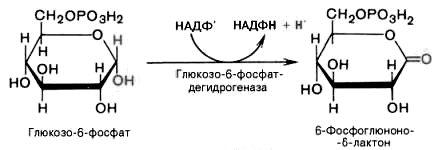
2).
6-фосфоглюконо--лактон
нестабилен и гидролизуется с образованием
6-фосфоглюконата (фермент –
6-фосфоглюконолактоназа):
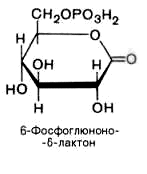
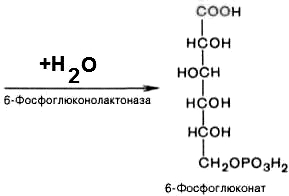
3).
Дегидрирование и декарбоксилирование
6-фосфоглюконата с образованием
рибулозо-5-фосфата (пентоза) и НАДФН при
участии декарбоксилирующей
6-фосфоглюконатдегидрогеназы:
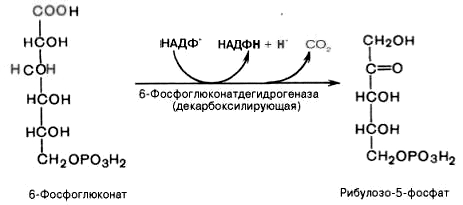
4).
Под действием эпимеразы из рибулозо-5-фосфата
образуется ксилулозо-5-фосфат (пентоза).
Под влиянием изомеразы рибулозо-5-фосфат
превращается в рибозо-5-фосфат (пентоза).
Между формами пентозофосфатов
устанавливается равновесие:
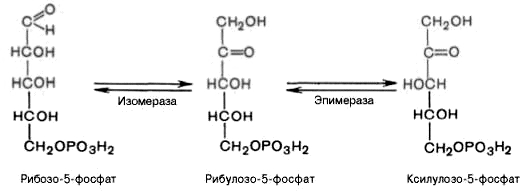
На
этом этапе пентозофосфатный путь может
быть завершен. При других условиях
наступает неокислительная стадия
пентозофосфатного цикла, протекающая
в анаэробных условиях. Она заключается
в переносе двух- и трехуглеродных
фрагментов от одной молекулы к другой.
При этом образуются вещества, характерные
для гликолиза (фруктозо-6-фосфат,
фруктозо-1,6-бисфосфат, фосфотриозы), и
вещества, специфические для пентозофосфатного
пути (седогептулозо-7-фосфат,
пентозо-5-фосфаты, эритрозо-4-фосфат).
Шесть
молекул
глюкозо-6-фосфата в пентозофосфатном
цикле
образуют 6 молекул
рибулозо-5-фосфата и 6 молекул
СО2.
Из 6 молекул
рибулозо-5-фосфата регенерируется 5
молекул
глюкозо-6-фосфата:
![]()
Промежуточные
продукты цикла (фруктозо-6-фосфат и
глицеральдегид-3-фосфат) включаются в
гликолиз.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник
Глюкоза
+ 2 АДФ + 2 H3PO4 =
2 Лактат + 2 АТФ + 2 Н2О
В
анаэробном гликолизе АТФ образуется
только за счет субстратного фосфорилирования
(2
моль АТФ на 1моль глюкозы).
Значение
анаэробного гликолиза особенно велико
для скелетных мышц. В интенсивно
работающих скелетных мышцах мощность
механизма транспорта кислорода
оказывается недостаточной для обеспечения
энергетической потребности, в этих
условиях резко усиливается анаэробный
гликолиз, и в мышцах накапливается
молочная кислота. Особое значение
анаэробный гликолиз имеет при
кратковременной интенсивной работе.
Так, бег в течение 30 с полностью
обеспечивается анаэробным гликолизом.
Однако уже через 4-5 мин бега энергия
поставляется поровну анаэробным и
аэробным процессами, а через 30 мин —
почти целиком аэробным процессом. При
длительной работе в аэробном процессе
используется не глюкоза, а жирные
кислоты.
Эритроциты
вообще не имеют митохондрий, поэтому
их потребность в АТФ удовлетворяется
только за счет анаэробного гликолиза.
Интенсивный гликолиз характерен также
для злокачественных опухолей.
Цикл Кори (глюкозо-лактатный цикл) и глюкозо-аланиновый цикл.
Молочная
кислота не является конечным продуктом
обмена, но ее образование — тупиковый
путь метаболизма: ее дальнейшее
использование связано опять с превращением
в пируват. Из клеток, в которых происходит
гликолиз, образующаяся молочная кислота
поступает в кровь и улавливается в
основном печенью, где и превращается в
пируват. Пируват в печени частично
окисляется, а частично превращается в
глюкозу. Часть пирувата в мышцах путем
трансаминирования превращается в
аланин, который транспортируется в
печень, и здесь снова образует пируват.
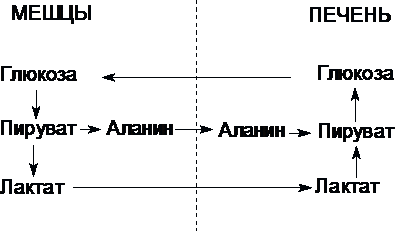
Рис.
Цикл Кори и глюкозо-аланиновый цикл
Биосинтез глюкозы (глюконеогенез).
Глюконеогенез
протекает в основном по тому же пути,
что и гликолиз, но в обратном направлении.
Однако три реакции гликолиза необратимы,
и на этих стадиях реакции глюконеогенеза
отличаются от реакций гликолиза.
Обходные пути глюконеогенеза.
I.
Первый обходной путь — превращение
пирувата в фосфоенолпируват.
Это
превращение осуществляется под действием
двух ферментов — пируваткарбоксилазы
(Е1) и карбокикиназы
фосфоенолпирувата (Е2).
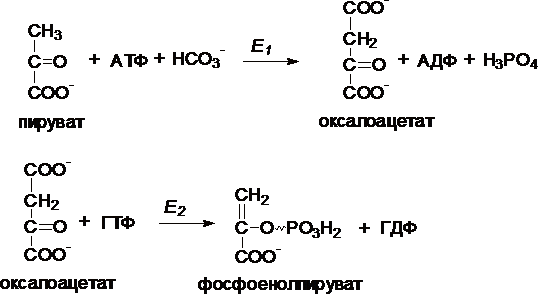
II. Превращение
фруктозо-1,6-дифосфата во
фруктозо-6-фосфат катализирует
фермент фруктозо-1,6-дифосфатаза.
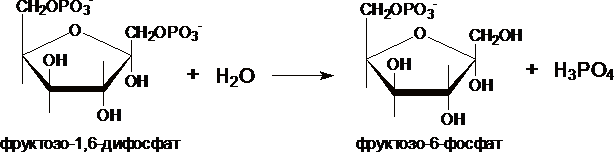
III. Образование
глюкозы из глюкозо-6-фосфата катализирует
фермент глюкозо-6-фосфотаза .
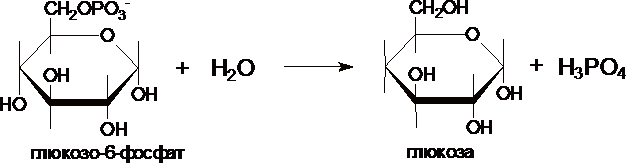
Этот
фермент отсутствует в клетках мозга,
скелетных мышц и др. тканях, поэтому
глюконеогенез, протекающий в этих тканях
не может поставлять глюкозу в кровь.
Глюкозо-6-фосфатаза локализована в
почках и, особенно, в печени, и
глюкозо-6-фосфат может превращаться в
глюкозу, если ее уровень в крови снижен.
Пентозофосфатный путь (пфп). Значение пфп.
Большая
часть глюкозы расщепляется по
гликолитическому пути. Наряду с гликолизом
существуют второстепенные пути
катаболизма глюкозы, одним из которых
является пентозофосфатный путь (ПФП).
Пентозофосфатный путь скорее даже —
анаболический путь, который использует
6 атомов С глюкозы для синтеза пентоз и
восстановительных эквивалентов в виде
НАДФН, необходимых для образования
липидов в организме. Однако, этот путь
— окисление глюкозы, и при определенных
условиях может завершиться полным
окислением глюкозы до СО2 и
Н2О.
Главные функции этого цикла следующие:
Генерирование
восстановительных эквивалентов в виде
НАДФН для восстановительных реакций
биосинтеза. Ферменты, которые катализируют
реакции восстановления, часто используют
в качестве кофактора НАДФ+/НАДФН,
а окислительные ферменты — НАД+/НАДН.
Восстановительные реакции биосинтеза
жирных кислот и стероидных гормонов
требуют НАДФН, поэтому клетки печени,
жировой ткани и др., где протекает синтез
липидов, имеют высокий уровень ферментов
ПФП. В печени около 30% глюкозы подвергаются
превращениям в
ПФП.
НАДФН
используется при обезвреживании лекарств
и чужеродных веществ в монооксигеназной
цепи окисления в печени.
НАДФН
предохраняет ненасыщенные жирные
кислоты эритроцитарной мембраны от
аномальных взаимодействий с активным
кислородом. Кроме того, эритроциты
используют реакции ПФП для генерирования
больших количеств НАДФН, который
необходим для восстановления глутатиона,
участвующего в системе обезвреживания
активного кислорода, что позволяет
поддерживать нормальную степень
окисления Fe2+ в
гемоглобине.
Снабжение
клеток рибозо-5-фосфатом для синтеза
нуклеотидов, нуклеотидных коферментов
и нуклеиновых кислот. Превращение
рибонуклеотидов в дезоксирибонуклеотиды
также требует НАДФН, поэтому многие
быстро пролиферирующие клетки требуют
больших количеств НАДФН.
Хотя
это не значительная функция ПФП, он
принимает участие в метаболизме пищевых
пентоз, входящих в состав нуклеиновых
кислот пищи и может превращать их
углеродный скелет в интермедиаты
гликолиза и глюконеогенеза.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
22.05.20158.12 Mб165Биохимия — краткий курс.pdf
- #
22.05.201515.29 Mб106Биохимия витаминов (с кафедры).djvu
- #
- #
- #
- #
- #
- #
- #
22.05.201511.34 Mб27Вазопатии. Геморрагический васкулит.Лейкозы Ч2.pdf
- #
- #
Источник
Лактат – конечный продукт анаэробного окисления глюкозы в мышцах, особенно в белых мышечных волокнах, где митохондрий меньше, чем в красных. Может включаться в глюконеогенез после окисления до пирувата в лактатдегидрогеназной реакции. При продолжительной физической работе основным источником лактата является скелетная мускулатура, в клетках которой преобладают анаэробные процессы. Накопление молочной кислоты в мышцах ограничивает их работоспособность. Это связано с тем, что при повышении концентрации молочной кислоты в ткани снижается уровень рН (молочнокислый ацидоз). Изменение рН приводит к ингибированию ферментов важнейших метаболических путей. В утилизации образующейся молочной кислоты важное место принадлежитглюкозо-лактатному циклу Кори.
Цикл Кори и глюкозо-аланиновый цикл (пояснения в тексте).
Лактат, образовавшийся в мышцах, переносится кровью в печень, где в процессе глюконеогенеза превращается в глюкозу, которая с током крови может возвращаться в работающую мышцу. В печени часть лактата может окисляться до углекислого газа и воды, превращаться в пируват и вовлекаться в общий путь катаболизма.
Значение цикла Кори:
1. Регуляция постоянного уровня глюкозы в крови.
2. Обеспечивает утилизацию лактата.
3. Предотвращает накопление лактата (снижение рН — лактоацидоз).
4. Экономичное использование углеводов организмом.
Регуляция обмена углеводов осуществляется на уровне тканей – кровь, печень, мышцы.
Глюкогенные аминокислоты, к которым относятся большинство белковых аминокислот. Ведущее место в глюконеогенезе среди аминокислот принадлежит аланину, который может превращаться в пируват путём трансаминирования. При голодании, физической работе и других состояниях в организме функционирует глюкозо-аланиновый цикл, подобный циклу Кори для лактата (рисунок 16.2). Существование цикла аланин – глюкоза препятствует отравлению организма, так как в мышцах нет ферментов, утилизирующих аммиак. В результате тренировки мощность этого цикла значительно возрастает.
Другие аминокислоты могут, подобно аланину, превращаться в пируват, а также в промежуточные продукты цикла Кребса (α-кетоглутарат, фумарат, сукцинил-КоА). Все эти метаболиты способны преобразовываться в оксалоацетат и включаться в глюконеогенез.
Глицерол – продукт гидролиза липидов в жировой ткани.Этот процесс значительно усиливается при голодании. В печени глицерол превращается в диоксиацетонфосфат – промежуточный продукт гликолиза и может быть использован в глюконеогенезе.
Жирные кислоты и ацетил-КоА не являются предшественниками глюкозы. Окисление этих соединений обеспечивает энергией процесс синтеза глюкозы.
Энергетический баланс. Путь синтеза глюкозы из пирувата (рисунок 16.6) содержит три реакции, сопровождающиеся потреблением энергии АТФ или ГТФ:
а) образование оксалоацетата из пирувата (затрачивается молекула АТФ);
б) образование фосфоенолпирувата из оксалоацетата (затрачивается молекула ГТФ);
в) обращение первого субстратного фосфорилирования – образование 1,3-дифосфоглицерата из 3-фосфоглицерата (затрачивается молекула АТФ).
Каждая из этих реакций повторяется дважды, так как для образования 1 молекулы глюкозы (С6) используются 2 молекулы пирувата (С3). Поэтому энергетический баланс синтеза глюкозы из пирувата составляет – 6 молекул нуклеозидтрифосфатов (4 молекулы АТФ и 2 молекулы ГТФ). При использовании других предшественников энергетический баланс биосинтеза глюкозы отличается.
Источник
10 фактов о молочной кислоте
- Лактат образуется всегда при производстве энергии.
Основной путь поступления энергии в клетки это деградация глюкозы. Молекула глюкозы подвергается серии из 10 последовательных реакций, чтобы получился пируват в ходе процесса называемого гликолиз. Далее одна часть пирувата частично окисляется и превращается в двуокись углерода и воду. Другая часть превращается в лактат под контролем фермента лактатдегидрогеназы. Эта реакция является обратимой.
- Часть лактата используется для синтеза энергии.
От 15 до 20 процентов от общего количества лактата превращается в гликоген в процессе глюконеогенеза.
- Лактат универсальный переносчик энергии.
В условиях высокого производства энергии в анаэробном режиме, лактат является переносчиком энергии из тех мест в которых невозможно провести трансформацию энергии, вследствие повышенной кислотности, в те места в которых она может быть трансформирована в энергию (сердце, дыхательные мышцы, медленно сокращающиеся мышечные волокна, другие группы мышц).
- Рост уровня лактата не является следствием недостатка кислорода.
Исследования на животных показывает, что внутриклеточный дефицит кислорода в изолированной мышце не показывает никаких ограничений активности дыхательной цепи митохондрий даже во время максимальной нагрузки. У нас всегда будет достаточно кислорода в мышцах.
- Лактат является индикатором нагрузки анаэробного гликолиза.
Каждый раз когда происходит образование пирувата, конечного продукта метаболизма глюкозы в процессе гликолиза, происходит образование лактата. Лактат накапливается просто потому, что скорости трансформации энергии в анаэробной и аэробной нагрузках отличаются.
- Чем быстрее бежит атлет-тем быстрее он производит лактат.
Уровень лактата в крови тесно связан с интенсивностью выполнения упражнения. Лактат накапливается из-за разницы скорости трансформации энергии в анаэробной и аэробной нагрузках. Скорость трансформации энергии при анаэробном метаболизме энергии быстрее чем при аэробном.
- Лактат не создает кислотность, а сопровождает ее.
Производя энергию мы одновременно производим кислотность. Энергетические реакции в нашем организме происходят при участии электронов как переносчиков энергии. Продуктами гликолиза являются лактат и протон водорода Н+. Мера активности (концентрация) ионов водорода (H+) в растворе выражает его кислотность.
Лактат только на время берет кислотного агента (Н+) для проведения реакции далее возвращая его в нейтральную среду.
- 90% лактата утилизируется организмом в первый час после тренировки.
60% лактата в организме полностью окисляется до СО2 и воды. Около 20% превращается в гликоген в процессе глюконеогенеза, часть используется для новообразования аминокислот. Лишь малая часть ( менее 5%) лактата выделяется с потом и мочой.
- Лактат в крови не будет систематически отражать присутствие лактата в мышцах.
Сравнения концентрации лактата в мышцах и крови показывают, что если усилие превышает 75-80% VO2max то концентрация лактата в мышцах (биопсия мышц передней поверхности бедра) выше чем в крови. В отличие от занятий умеренной интенсивности 30%,50%,70% VO2max где концентрация лактата в артериальной крови выше чем в мышцах.
- Лактат не вызывает боль и судороги в мышцах.
Болезненные ощущения в мышцах на следующий день после интенсивной тренировки вызваны повреждениями мышц и воспалением тканей, которые происходят после выполнения упражнения. Большинство мышечных судорог вызывается нервными рецепторами мышц, которые перевозбуждаются с появлением усталости в мышцах.
Автор: Игорь Лашкевич
Диссоциация молочной кислоты
Рис. 2.1. Зависимость между степенью диссоциации молочной кислоты и значением pH. Вы видите, как меняется pH при изменении отношения лактат/молочная кислота. Когда концентрации лактата и молочной кислоты одинаковы (т.е. значение дроби равняется 1), pH равен значению рК для молочной кислоты (таким образом, рК молочной кислоты = 3,85)
На рис. 2.1 показано, как изменяется отношение концентраций лактата и молочной кислоты с увеличением pH. Когда концентрации лактата и молочной кислоты равны (т.е. отношение лактат/молочная кислота =1), pH принимает значение рК для молочной кислоты, равной 3,85.
Молочная кислота и бикарбонатная буферная система
Рис. 2.2. Гомеостаз молочной кислоты и pH поддерживается бикарбонатной буферной системой. Бикарбонатная буферная система удаляет из раствора протоны [Н+], образующиеся в процессе анаэробного гликолиза. Протоны связываются при образовании молекулы воды, а СO2 выводится через легкие
Чтобы продемонстрировать работу бикарбонатной буферной системы в домашних условиях in vivo, понадобится всего несколько минут. Просто обеспечьте себе физическую нагрузку в анаэробных условиях: пробегитесь как можно быстрее (лучше в гору), задержав при этом дыхание. В это время в ваших мышцах будет проходить анаэробный гликолиз с образованием молочной кислоты, которая диссоциирует на лактат и протон [Н+][1]. Чтобы снизить концентрацию протонов, с ними реагирует бикарбонат, а образующаяся угольная кислота распадается на воду и СO2. Повышение концентрации СO2 стимулирует гипервентиляцию легких, и избыток образованного С02 удаляется вместе с выдыхаемым воздухом.
Метаболизм лактата: цикл Кори
Лактат постоянно образуется из глюкозы в процессе анаэробного гликолиза в эритроцитах, сетчатке и мозговом веществе почек. Этот лактат снова превращается в глюкозу в цикле Кори. Лактат переносится в печень и превращается в глюкозу в процессе глюконеогенеза. Образование глюкозы из лактата требует затраты 6 молекул АТФ. Если
из-за поражения печени цикл Кори блокируется, в организме накапливается лактат и развивается гиперлактатемия. Гиперлактатемия часто протекает бессимптомно, и это весьма распространенное состояние, не представляющее угрозы для жизни. Лишь в редких случаях развивается лактацидоз, с которым не справляются буферные системы организма.
БИОЛОГИЧЕСКИЙ КОНТРОЛЬ (МОНИТОРИНГ) В СПОРТИВНОЙ ТРЕНИРОВКЕ
Источник:
Учебное пособие для ВУЗов «Спортивная физиология».
Автор: И.И. Земцова Изд.: Олимпийская лит-ра, 2010 год.
Общей целью биологического контроля в спорте является повышение эффективности спортивной тренировки за счет оптимизации физической нагрузки на основе объективной оценки функциональной подготовленности спортсмена.
На разных этапах подготовки спортсменов стоят разные задачи, в соответствии с которыми определяют цель и формы контроля. В теории и практике спорта различают четыре основных вида контроля: оперативный, текущий, этапный и углубленный (Волков, 1996; Биологический контроль…, 1996; Куроченко, 2005; Левушкин, 2001; Платонов, 1997; Clausen, 1997).
Оперативный контроль (срочный) предполагает оценку оперативных состояний — срочных реакций организма спортсменов на нагрузку в ходе отдельных тренировочных занятий и соревнований.
Текущий контроль направлен на оценку текущих состояний, являющихся следствием физических нагрузок серии занятий, тренировочных или соревновательных микроциклов.
Этапный контроль позволяет оценить состояние спортсмена, являющееся следствием долговременного тренировочного эффекта на определенных этапах подготовки.
Углубленный контроль проводят один раз в году для комплексной оценки подготовленности спортсмена и состояния его здоровья.
Показатели, используемые соответственно определенному виду контроля, должны быть информативными и надежными, соответствовать:
- специфике вида спорта;
- возрасту и квалификации испытуемых;
- направленности тренировочного процесса.
В видах спорта, связанных с проявлением выносливости (плавание, гребля, велосипедный спорт, лыжные гонки, бег на средние и длинные дистанции и др.), преимущественно исследуют показатели, характеризующие состояние сердечно-сосудистой и дыхательной систем, обменных процессов. Благодаря им можно наиболее достоверно оценить потенциальные возможности спортсменов в достижении высоких спортивных результатов.
В скоростно-силовых видах спорта, где главной задачей является умение проявлять кратковременные мышечные напряжения (спринтерский бег, легкоатлетические прыжки и метания, тяжелая атлетика, отдельные дисциплины велосипедного, конькобежного спорта, плавания и др.) как средства контроля используют показатели, характеризующие состояние нервно-мышечного аппарата, ЦНС, скоростно-силовых компонентов двигательной функции, которые проявляются в специфических тестовых упражнениях.
В видах спорта, где спортивные достижения преимущественно обусловлены деятельностью анализаторов, подвижностью нервных процессов, которые обеспечивают точность, размеренность движений во времени и пространстве (гимнастика, акробатика, фигурное катание, прыжки в воду, все виды спортивных игр, стрельба и др.), в процессе контроля используют широкий комплекс показателей. Они характеризуют точность воспроизведения временных, пространственных и силовых параметров специфических движений, способность к обработке информации и быстрому принятию решений, эластичность скелетных мышц, подвижность суставов, координационные возможности и др. (Белоцерковский, 2005; Биологический контроль…, 1996; Бргсюн, 2003; Платонов, 1997).
Использование содержания лактата в крови в процессе контроля за подготовкой спортсменов
Источник:
Учебное пособие для ВУЗов «Спортивная физиология».
Автор: И.И. Земцова Изд.: Олимпийская лит-ра, 2010 год.
Молочная кислота образуется только в анаэробных условиях. Большое ее количество вызывает изменения концентрации ионов гидрогена во внутренней среде организма. При значительном смещении pH в сторону повышения кислотности наблюдается угнетение активности ферментов, регулирующих способность мышц к сокращению и скорость анаэробного ресинтеза АТФ (АТФаза миофибрилл, креатинфосфокиниза, ферменты гликолиза) (Буланов, 2002; Волков и соавт., 1998; Ключевые факторы адаптации…, 1996; Мохан, Глессон, Грингафор, 2001; Тнимова, 2004).
Накопление молочной кислоты в саркоплазматическом пространстве мышц сопровождается изменением осмотического давления, что ведет к поступлению воды с межклеточной среды внутрь мышечных волокон, вызывая их набухание. Значительные изменения осмотического давления в мышцах могут служить причиной болевых ощущений, так как набухшие клетки сдавливают нервные окончания.
Поступая из работающих мышц в кровь, молочная кислота взаимодействует с гидрокарбонатной буферной системой, что вызывает выделение «не метаболического» избытка СO2, следствием чего является ускорение ЛВ.
Молочная кислота и лактат — не одно и то же соединение. Лактат — это соль молочной кислоты, образующаяся при замещении Н+ молочной кислоты на Na+ или К+. В результате анаэробного гликолиза образуется молочная кислота, которая очень быстро превращается в соль—лактат. Поэтому, говоря об этом соединении в крови, целесообразно использовать термин «лактат».
Если спортсмен находится в состоянии покоя, содержание лактата в его крови составляет 1,0—2,5 г*лг. В первые 2—19-ю минуты работы содержание лактата быстро увеличивается, а затем стабилизируется. В случае выполнения работы с кислородным запросом более 80 % содержание лактата в крови постоянно увеличивается, достигая максимальных значений не во время работы, а на 2—10-й минутах восстановительного периода.
Содержание лактата в крови довольно точно характеризует направленность тренировочных занятий, и поэтому определение его содержания в процессе занятий — один из важнейших методов оперативного управления нагрузкой.
Определены нормативные показатели зон работы разной интенсивности по изменению уровня лактата в крови в зависимости от мощности работы (Платонов, 1995; Лактатный порог…, 1997):
1-я зона — аэробно-восстановительная работа, направленная на устранение недоокисленных продуктов обмена веществ, образовавшихся во время предыдущей работы. В этом случае смещения биохимических показателей крови находятся в пределах нормы. Концентрация лактата составляет от 1 до 3 ммоль-л-1, ЧСС — до 130 уд мин-1;
2-я зона — аэробно-тренировочная работа, направленная на стабилизацию работоспособности. Концентрация лактата в крови составляет 3,5— 4,5 ммоль-л-1, ЧСС — 130—150 уд-мин-1;
3-я зона — смешанная работа аэробной направленности, проводится с целью повышения уровня VO2max. Концентрация лактата в крови составляет от 4,5 до 7,5 ммоль-л-1, ЧСС — 150—170 уд-мин-1;
4-я зона — смешанная работа анаэробной направленности, проводится для развития выносливости. Концентрация лактата в крови 8—11 ммоль-л-1, ЧСС — 170—190 уд мин-1;
5-я зона — соревновательно-специфическая работа, направленная на развитие скоростных качеств. При этом наблюдается изменение биохимических показателей крови до индивидуального максимума. Концентрация лактата может достигать 28 ммоль-л-1, а ЧСС — индивидуального максимума.
Если после проведения тренировки, направленной на развитие аэробной работоспособности, концентрация лактата в крови меньше 4 ммоль-л-1, это значит, что интенсивность его недостаточна. Тренировочные занятия, направленные на развитие силовой выносливости, должны осуществляться при содержании лактата в крови 5—6 ммоль-л-1; занятия, направленные на экономизацию анаэробного обмена, — 8—11 ммоль-л-1.
Показатель лактата в крови кроме использования для нужд оперативного контроля может дать информацию о мощности и емкости гликолитического механизма энергообеспечения, о состоянии тренированности:
Реакция | Оценка тренированности |
Уменьшение содержания лактата в крови при стандартной физической нагрузке | Повышение тренированности |
Повышение содержания лактата в крови при повышении мощности тестирующей нагрузки | Совершенствование анаэробных процессов энергообеспечения работы |
Отсутствие изменений содержания лактата или его уменьшение при повышении мощности тестирующей нагрузки | Рост экономизации функций организма (рост тренированности) |
Отсутствие изменений содержания лактата в крови при снижении мощности тестирующей нагрузки | Снижение тренированности |
Резкое увеличение содержания лактата в крови при сохранении постоянной мощности работы | Низкая тренированность |
Этот показатель может использоваться для определения лактатного порога, протекания восстановительных процессов, если уровень лактата в крови определять в динамике после нагрузок различного характера (Евгеньева, 2002; Лактатный порог…, 1997; Симонова, 2001; Шац, 2001).
Оснащение: лактометр с тестовыми полосками или набор реактивов для определения лактата в крови (например, фирмы Dr. Lange, Германия), фотометр этой же фирмы, микропипетка и капилляры для забора крови, скарификаторы, спирт, вата.
Ход работы
В состоянии покоя у испытуемых (желательно с разным уровнем тренированности) берут по 10 мкл крови из пальца и вносят в бутылочки с готовым реактивом.
Испытуемые в условиях стадиона выполняют бег на 800 м с регистрацией времени преодоления этой дистанции, или в условиях лаборатории — бег на месте в течение 3 мин с регистрацией количества шагов. После бега, на 3-й и 7-й минутах восстановления, у испытуемых берут по 10 мкл крови из пальца, вносят в бутылочку с реактивом и определяют содержание лактата в крови на фотометре фирмы Dr. Lange (Германия) при длине волны 420 нм по инструкции, прилагаемой к прибору.
Полученные результаты вносят в таблицу 54, сравнивают данные, полученные во время обследования двух испытуемых, с приведенными выше, делают выводы о гликолитической мощности испытуемых, состоянии их тренированности.
Таблица 54 — Содержание лактата в крови (ммоль-л-1) после выполнения физической нагрузки субмаксимальной интенсивности и в процессе восстановления после нее
Испытуемый | Лактат в крови Связанные материалы | |||