Шари кори головного мозку
Взаємовідносини борозен і звивини з кістками і швами черепа у новонародженої дитини інші, ніж у дорослого. Основні борозни (центральна, латеральна) виражені добре, але гілки основних борозен і дрібні звивини позначені слабо. Надалі в процесі розвитку кори борозни стають глибшими, а звивини між ними рельєфніше. Співвідношення борозен, звивин і швів черепа, характерне для дорослої людини, встановлюється у дітей в 6-8 років.
Протягом перших місяців життя розвиток кори йде дуже швидкими темпами. Більшість нейронів набувають зрілу форму, інтенсивно відбуваються процеси мієлінізації нервових волокон, що дозволяють реагувати на зовнішні подразники більш диференційовано.
У процесі еволюції людини як біологічного виду, а також у процесі онтогенезу — індивідуального розвитку кожної людини — відбувається кортікалізаціі функцій, тобто включення кори головного мозку в регуляцію функцій нижележащих структур мозку. Це дозволяє організувати більш досконалу, що враховує індивідуальний досвід, збережений у пам’яті, регуляцію функцій організму. Надалі, у міру автоматизації тієї чи іншої реакції, виконання її знову передається підкірковим структурам з формуванням автоматичного реагування.
Різні коркові зони дозрівають нерівномірно. Найбільш рано дозріває соматосенсорная і рухова кора, дещо пізніше — зорова і слухова. Особливо інтенсивним протягом першого півріччя життя є розвиток зорової кори, що тягне за собою розвиток інших зон мозку та їх інтеграцію. Дозрівання сенсорних і моторних зон в основному закінчується до 3 років. Значно пізніше дозріває асоціативна кора: до 7 років формуються її основні зв’язки, а остаточна диференціювання, формування нейронних ансамблів і зв`язків з іншими відділами мозку відбуваються до підліткового віку. Найбільш пізно (ближче до 9 років) дозрівають лобні області кори. Поступовість дозрівання структур кори великих півкуль визначає вікові особливості вищих нервових функцій і поведінкових реакцій дітей різних вікових груп.
Загальна площа кори головного мозку людини близько 2200 см2, число нейронів кори перевищує 10 млрд. У складі кори маються пірамідні, зірчасті, веретеноподібні нейрони.
Пірамідні нейрони мають різну величину, аксон пірамідного нейрона, як правило, проходить через білу речовину в інші зони кори або в інші мозкові структури.
Зірчасті клітини мають короткі добре гілкуються дендрити і короткий аксон, що забезпечує зв’язки нейронів в межах самої кори головного мозку.
Веретеноподібні нейрони забезпечують вертикальні або горизонтальні взаємозв’язку нейронів різних шарів кори.
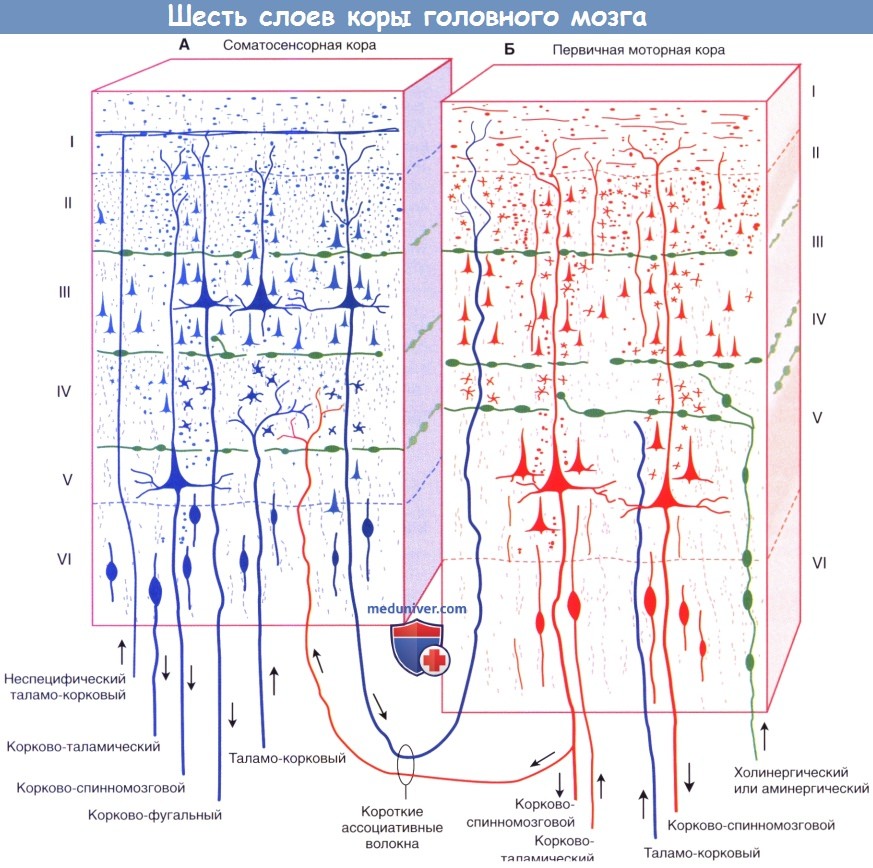
Кора головного мозку має переважно шестіслойних будову (рис. 11.10).

Рис. 11.10. Будова кори головного мозку
Шар I — верхній молекулярний, представлений в основному ветвлениями висхідних дендритів пірамідних нейронів, серед яких розташовані рідкісні горизонтальні клітини і клітини-зерна, сюди ж приходять волокна неспецифічних ядер таламуса, що регулюють через дендрити цього шару рівень збудливості кори головного мозку.
Шар II — зовнішній зернистий, складається з зірчастих клітин, що визначають тривалість циркулювання збудження в корі головного мозку, тобто мають відношення до пам’яті.
Шар III — зовнішній пірамідний, формується з пірамідних клітин малої величини і разом з II шаром забезпечує Корко-коркові зв’язку різних звивин мозку.
Шар IV — внутрішній зернистий, містить переважно зірчасті клітини. Тут закінчуються специфічні таламокортікальние шляху, тобто шляху, що починаються від рецепторів аналізаторів.
Шар V — внутрішній пірамідний (гангліозний), шар великих пірамід, які є вихідними нейронами, аксони їх йдуть в стовбур мозку і спинний мозок. У руховій зоні в цьому шарі знаходяться гігантські пірамідні клітини, відкриті Бецом (клітини Беца).
Шар VI — шар поліморфних клітин, більшість нейронів цього шару утворюють кортико-таламические шляху.
Розподіл нейронів по шарах в різних областях кори дозволило виділити в мозку людини 53 цитоархитектонических поля (поля Бродмана), які вдосконалюються в міру розвитку кори головного мозку. У людини і вищих ссавців розрізняють, поряд з первинними, вторинні і третинні коркові поля, що забезпечують асоціацію функцій даного аналізатора з функціями інших аналізаторів.
Особливістю коркових полів є екранний принцип їх функціонування, що полягає в тому, що рецептор проектує свій сигнал не так на один нейрон кори, а на поле нейронів, яке утворюється їх зв’язками. У результаті сигнал фокусується не крапка в точку, а на безлічі різноманітних нейронів, що забезпечує його повний аналіз і можливість передачі в інші зацікавлені структури. Так, одне волокно, що приходить в зорову область кори, може активувати зону розміром 0,1 мм. Це означає, що один аксон розподіляє свою дію на більш ніж 5000 нейронів.
Функції окремих зон нової кори визначаються особливостями її структурної організації, зв’язками з іншими структурами мозку, участю в сприйнятті, зберіганні та відтворенні інформації при організації та реалізації поведінки, регуляції функцій сенсорних систем, внутрішніх органів.
Структурний відмінність ділянок кори головного мозку пов’язане з різницею їхніх функцій. У корі головного мозку виділяють сенсорні, моторні і асоціативні області (рис. 11.11).

Рис. 11.11. Цітоархітектоніческі поля Бродмана
Коркові кінці аналізаторів мають свою топографію — локальне розташування в певних ділянках кори головного мозку. Вони називаються сенсорними областями кори головного мозку. Коркові кінці аналізаторів різних сенсорних систем перекриваються. Крім цього, у кожній сенсорній системі кори маються полісенсорній нейрони, які реагують не тільки на «свій» адекватний стимул, але й на сигнали інших сенсорних систем. Ці механізми лежать в основі формування полімодальних зв’язків, що забезпечують сочетанную реакцію на різні подразники.
Шкірна рецептірующая система, таламокортікальние шляху проектуються на задню центральну звивину. Тут мається суворе соматотопическую поділ. На верхні відділи цієї звивини проектуються рецептивні поля шкіри нижніх кінцівок, на середні — тулуба, на нижні відділи — руки, голови.
На задню центральну звивину в основному проектується больова і температурна чутливість. У корі тім’яної частки (поля 5 і 7, див. Рис. 11.11), де також закінчуються провідні шляхи чутливості, здійснюється більш складний аналіз: локалізація роздратування, дискримінація, стереогіоз. При пошкодженнях кори особливо сильно порушуються функції дистальних відділів кінцівок, особливо рук.
Зорова система знаходиться в потиличній частці мозку: поля 17, 18, 19. Центральний зоровий шлях закінчується в поле 17; він інформує про наявність та інтенсивності зорового сигналу. У полях 18 і 19 аналізуються колір, форма, розміри, якість предметів. Поразка поля 19 кори головного мозку призводить до того, що хворий бачить, але не впізнає предмет (зорова агнозія, при цьому втрачається також колірна пам’ять).
Слухова система проектується в поперечних скроневих звивинах (звивини Гешля), в глибині задніх відділів латеральної (Сільвієвій) борозни (поля 41, 42, 52). Саме тут закінчуються аксони задніх горбів Четверохолміе і латеральних колінчастих тіл.
Нюхова система проектується в області переднього кінця гіппокампальних звивини (поле 34). Кора цій області має не шести-, а тришарове будову. При подразненні відзначаються нюхові галюцинації, пошкодження її веде до аносмія (втрати нюху).
Смакова система проектується в гіппокампальних звивині по сусідству з нюхової областю кори (поле 43).
У передній центральній звивині розташовані зони, подразнення яких викликає рух, вони представлені по соматотопическую типу, але зовсім інакше: у верхніх відділах звивини — нижні кінцівки, в нижніх — верхні. Це моторні області кори головного мозку.
Спереду від передньої центральної звивини лежать премоторні поля 6 і 8. Вони організовують не ізольовані, а комплексні, координовані, стереотипні рухи. Ці поля також забезпечують регуляцію тонусу гладкої мускулатури, пластичний тонус м’язів через підкіркові структури.
У реалізації моторних функцій беруть участь також друга лобова звивина, потилична, верхнетеменная області.
Рухова область кори як ніяка інша має велику кількість зв’язків з іншими аналізаторами, чим, мабуть, і зумовлена наявність в пий значного числа полісенсорних нейронів.
Всі сенсорні проекційні зони і моторні області кори займають менше 20% поверхні кори головного мозку. Решта — це асоціативні області. Кожна асоціативна область кори пов’язана потужними зв’язками з кількома проекційними областями. У асоціативних областях відбувається інтеграція разномодальних інформації, що дозволяє здійснювати усвідомлення надходить інформації та складні поведінкові акти. Асоціативні області мозку у людини найбільш виражені в лобової, тім’яної і скроневій.
Кожна проекційна область кори оточена асоціативними областями. Нейрони цих областей здатні до сприйняття разномодальних інформації, володіють великими здібностями до навчання. Полісенсорній нейронів асоціативної області кори забезпечує їх участь в об’єднанні, що надходить, забезпеченні взаємодії сенсорних і моторних областей кори.
Так, в тім’яній асоціативної області кори формуються суб’єктивні уявлення про навколишній простір, про нашому тілі. Це стає можливим завдяки зіставленню соматосенсорной, пропріоцептивної і зорової інформації. Лобові асоціативні поля мають зв’язку з лимбичним відділом мозку і беруть участь в організації програм дії при реалізації складних поведінкових актів з урахуванням їх емоційного забарвлення.
Першою і найбільш характерною рисою асоціативних областей кори є здатність їх нейронів сприймати разномодальную інформацію, причому сюди надходить не первинна, а вже оброблена інформація з виділенням біологічної значущості сигналу. Це дозволяє формувати програму цілеспрямованої поведінкового акту.
Друга особливість асоціативної області кори полягає в здатності до пластичних перебудовам в залежності від значимості надходить.
Третя особливість асоціативної області кори виявляється у тривалому зберіганні слідів сенсорних впливів. Руйнування асоціативної області призводить до виражених порушень навчання, пам’яті.
Розподіл функцій по областях мозку не є абсолютним. Встановлено, що практично всі області мозку мають полісенсорній нейрони, які певною мірою можуть брати на себе функцію пошкоджених модальноспсціфічсскіх нейронів. Це дозволяє компенсувати пошкодження структур мозку в ті періоди дитинства, коли пошкоджена функція ще не закріплена жорстко в структурі нервової тканини.
Важливою особливістю кори головного мозку є її здатність тривало зберігати сліди збудження. Це властивість надає корі виняткове значення в механізмах асоціативної переробки та зберігання інформації, накопичення знань.
Источник
Строение коры головного мозга: слои, клеткиТолщина коры больших полушарий (от греч. pallium — мантия) варьирует от 2 до 4 мм, наиболее тонкая ее часть находится в области первичной зрительной коры, а наиболее утолщенная — в первичной моторной области. Больше половины общей поверхности коры скрыто стенками борозд. Мозг содержит приблизительно 86 млрд, нейронов (кора больших полушарий содержит только 19% общего количества, но составляет 81% массы мозга), аналогичное число глиальных клеток и разветвленную капиллярную сеть. Микроскопически кора имеет полосчатый, или пластинчатый, вид, отражающий распределение ее клеток и нервных волокон, а также веерообразную организацию клеточных элементов. Общая цитоархитектоника (схема строения, основанная на типе клеток; схему строения в зависимости от распределения миелинизированных волокон обозначают термином миелоархитектоника) во многом варьирует от зоны к зоне, что позволяет составить «карту» коры с десятками гистологически различных зон. Больших успехов удалось достичь в соотнесении этих зон со специфическими функциями. Несмотря на то, что это имеет теоретическую ценность, подобное разделение довольно упрощено, так как часто принимают во внимание только ключевые функции более обширных функциональных систем, взаимодействующих с другими отделами мозга. а) Пластинчатое строение. Пластинчатое распределение нейронов характерно для всех отделов коры. Филогенетически «старые отделы», включая древнюю кору (обонятельную кору) и старую кору (гиппокампальная формация и зубчатое ядро; участвуют в процессах памяти) образованы тремя клеточными пластинками. Однако в новой коре (неокортексе или изокортексе, название которой указывает на единый нейрогенез коры, закончившийся образованием шестислойного строения), составляющей оставшиеся 90% (подавляющую часть коры больших полушарий), происходит переход к шестислойному строению.
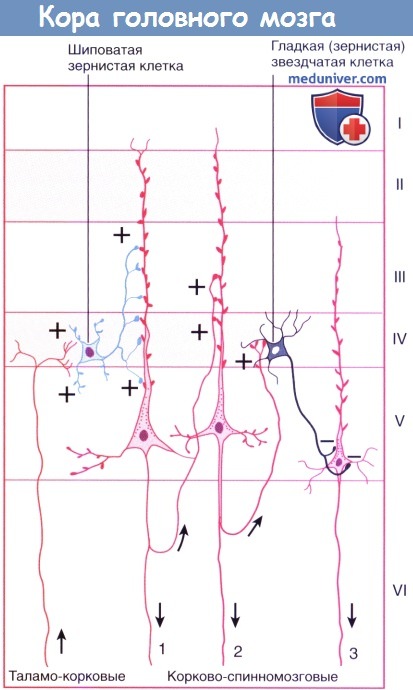
Клеточные пластинки новой коры (неокортекса): I. Молекулярный слой образован концами апикальных дендритов пирамидных клеток и наиболее дистальными ветвями аксонов, проходящих к коре от интраламинарных ядер таламуса. II. Наружный зернистый слой образован мелкими пирамидными и звездчатыми клетками. III. Наружный пирамидный слой состоит из средних пирамидных клеток и звездчатых клеток. IV. Внутренний зернистый слой образован звездчатыми клетками, к которым подходят восходящие волокна от релейных таламических ядер. Звездчатые (зернистые) клетки особенно многочисленны в первичной соматосенсорной коре, первичной зрительной коре и первичной слуховой коре; к ним подходят афферентные чувствительные волокна. Эти отделы обозначают как гранулярная кора. Наоборот, первичная моторная кора — место начала корково-спинномозгового и корково-бульбарного путей, имеет относительно небольшое количество зернистых клеток в слое IV и большое число пирамидных клеток в слоях III и V, которые смазывают (перекрывают) отдельные слои. Эту область обозначают как агранулярная кора. V. Внутренний пирамидный слой образован крупными пирамидными клетками, волокна которых направляются к полосатому телу, стволу мозга и спинному мозгу. VI. Слой веретеновидных клеток образован видоизмененными пирамидными клетками, волокна которых направляются к таламусу. б) Колончатое строение. Помимо хорошо различимой пластинчатой организации коры больших полушарий, можно наблюдать также лучистую, или «колончатую», организацию клеточных структур. Колончатое строение новой коры стало основой исследования функционирования групп нейронов в соматосенсорной коре животных. Подобные лучевидно расположенные группы клеток, вероятно, представляют собой отдельные зоны с одинаковыми физиологическими свойствами и образуют структурную основу для создания более сложных функций. Группы колонн могут образовывать модули, участвующие в обработке различных сенсорных модальностей и функций. Установлено, что колонны в различных отделах коры имеют разное строение, обусловленное вариабельностью многочисленных свойств, включая строение и число клеток, онтогенез, синаптические связи и молекулярные маркеры. Все это влияет на различные функциональные характеристики и реакции в ответ на возбуждение. В качестве организующего принципа подобная концепция колончатого строения представляет определенную пользу, однако удобнее рассматривать кору как совокупность горизонтального (пластинчатого) и вертикального (лучистого) типов строения. Несмотря на то, что колонны не представляют собой отдельные подобные образования с четкими границами, такая концепция более точно отражает строение коры, а также наблюдаемые экспериментально функциональность, «экономию пространства» и пластичность коры. Взаимодействие между группами колонн позволяет осуществлять более сложную деятельность, модели поведения или решать когнитивные задачи. Лежащая в основе организации коры «схема» приводит к тому, что каждая из колонн становится модально (функционально) специфичной после «обработки» информации отдельными элементами. Однако конечная реакция проекционных нейронов колонн может значительно варьировать в зависимости от степени их возбуждения и информации, подходящей к каждому нейрону. Например, одна колонна может реагировать на движение в определенном суставе, но не на раздражение кожи над ним; однако при изменении условий их реакция также может изменяться.
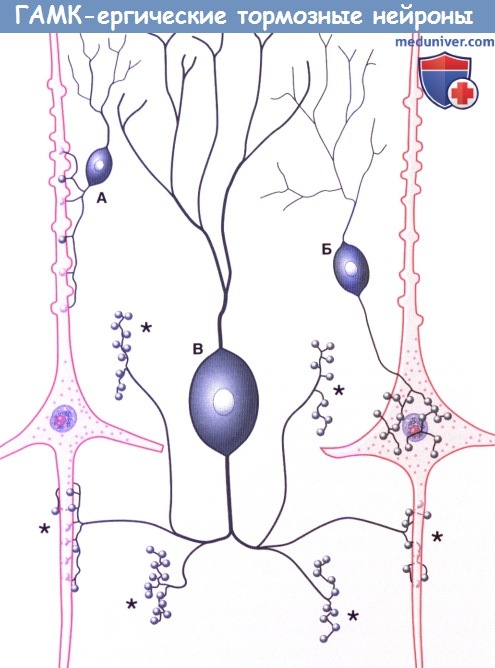
в) Типы клеток. Морфологически кортикальные нейроны разделяют на две большие группы. Большая часть (60-85%) представлена пирамидными нейронами (из-за их формы) — единственными, волокна которых покидают кору (и к которым подходит большая часть волокон), что объясняет их альтернативное название — корковые проекционные нейроны; их волокна — возбуждающие глутаматергические. К оставшимся 15-40% нейронам относят непирамидные, или вставочные, нейроны; несмотря на то, их взаимосвязи не выходят за пределы коры, они осуществляют регуляцию и значительное влияние на ее деятельность; их тип — преимущественно тормозной ГАМК-ергический. Внутри каждой группы можно выделить множество подгрупп в зависимости от морфологии, взаимосвязей, электрофизиологических свойств, типа развития, физиологических характеристик, молекулярных маркеров и т. д. (Примерами основных морфологических и функциональных клеточных типов служат пирамидные клетки, шиповатые зернистые клетки (измененные пирамидные клетки) и группа непирамидных тормозных вставочных нейронов.) • Пирамидные клетки имеют пирамидоподобную форму с вершиной, направленной к поверхности. Размеры клеток в высоту составляют 20-30 нм в слоях II и III и более чем в два раза больше в слое V. Самые крупные — гигантские пирамидные клетки Беца — расположены в моторной коре. Единственный апикальный дендрит каждой пирамидной клетки достигает слоя I, заканчиваясь на пучке дендритов. Несколько базальных дендритных ветвей отходят от базальных «углов» клетки и веерообразно расходятся к соответствующим слоям. Апикальные и базальные дендритные ветви свободно разветвляются и усыпаны дендритными шипиками. Большинство пирамидных клеток расположено в слоях II-III и V-VI. Отходящие от основания клеток аксоны отдают несколько возвратных ветвей до вхождения в подлежащее белое вещество, их функция — возбуждение соседних пирамидных клеток. • Шиповатые зернистые клетки — один из вариантов атипичных пирамидных клеток, лежащих в слое IV и наиболее многочисленных в первичной сенсорной коре. Их шиповатые дендриты не выходят за пределы слоя IV, а аксоны могут подниматься или опускаться, образуя возбуждающие глутаматергические синаптические контакты с пирамидными клетками. На них переключается большая часть волокон восходящих таламических путей к слою IV с дальнейшим радиальным распространением. Нейромедиатором различных по структуре и классификации непирамидных тормозных вставочных нейронов служит ГАМК. [Сложная классификация нейронов новой коры постоянно меняется. Гладкие звездчатые (зернистые) клетки можно обнаружить во всех слоях коры; их дендриты веерообразно расходятся во всех направлениях, а их аксоны образуют локальные древовидные разветвления, поэтому их иногда называют нейронами локальных сплетений. Несмотря на их уникальные морфологические характеристики, нейроглиеформные, клетки-канделябры и корзинчатые клетки считают специализированными типами зернистых клеток. Наш совет: если Вы встречаете термины «зернистая» или «гладкая звездчатая» клетка, для облегчения чтения и понимания их следует рассматривать как вставочные нейроны.] Для организации их можно разделить на три крупных семейства в зависимости от экспрессируемых этими вставочными нейронами биомаркеров: парвальбумин, соматостатин и серотонин (5-гидрокситриптамин, 5НТ) 3а-рецептор (5HT3aR). • Парвальбумин-экспрессирующие вставочные нейроны не имеют шигшков на дендритах. К ним подходят возбуждающие волокна от таламуса и коры, а тормозные — от других вставочных нейронов того же типа. Считают, что они играют роль в стабилизации активности корковых нейронных сетей. Как и в коре мозжечка, эти нейроны обладают фокусирующим действием на кору больших полушарий, тормозя слабо возбужденные колонны клеток. Клетки-канделябры (названы так из-за канделяброподобных групп аксоаксональных синаптических окончаний) наиболее многочисленны в слое II, образуют контакты с начальным сегментом аксона пирамидной клетки и играют ключевую роль в корково-корковых взаимодействиях. Корзинчатые клетки лежат преимущественно в слоях II и V, а свое название получили благодаря тому, что их аксоны формируют околоклеточные «корзинки» вокруг тел пирамидных клеток, дистальных отделов их дендритов и аксонов других корзинчатых клеток. • Соматостатин-экспрессирующие вставочные нейроны. Примером служат клетки Мартинотти, расположенные в пластинках V и VI, аксоны которых направляются к пластинке I. Получая сигналы от пирамидных клеток, они способны ограничивать их периферическое возбуждение и объединять несенсорную информацию, осуществляя, таким образом, регулирование обработки сигналов в дендритах их пирамидных клеток в зависимости от внешних обстоятельств. • 5НТ3а-экспрессирующие вставочные нейроны — гетерогенная группа, к которой относят большую часть вставочных нейронов поверхностных корковых слоев. Получая информацию от корковых и таламических нейронов, они могут участвовать в процессах обучения, осуществляя влияние на корковые нейронные цепи. Он нейроглиеформных клеток (паутинообразных клеток)—одного из главных типов вставочных нейронов слоев II и III — веерообразно расходятся дендриты, обладающие уникальным свойством образования синапсов друг с другом и другими типами вставочных нейронов; этот факт указывает на их важнейшую роль в синхронизации корковых нейронных цепей. Другая морфологически гетерогенная группа вставочных нейронов, помимо ГАМК, экспрессирует вазоактивный интестинальный полипептид; другие типы вставочных нейронов этой группы экспрессируют также холецистокинин и другие пептидные рецепторы.
г) Афферентные волокна. Восходящие волокна к любому отделу коры могут иметь четыре различных источника (в первую очередь, корковые) и оканчиваются в разных отделах. 1. Длинные и короткие ассоциативные волокна от мелких и средних пирамидных клеток в слоях II и III из других корковых зон того же полушария. 2. Комиссуральные волокна от средних пирамидных клеток слоев II и III проходят через мозолистое тело от аналогичных или топографически идентичных участков противоположного полушария. 3. Таламо-корковые волокна от соответствующего специфического или ассоциативного ядра (например, волокна от вентрального заднего таламического ядра к соматосенсорной коре) и от дорсомедиального таламического ядра к предлобной (префронтальной) коре (описана ниже) оканчиваются в пластинке IV. Неспецифические таламо-корковые волокна от внутрипластинчатых ядер оканчиваются во всех пластинках. 4. Холинернические и аминергические волокна от базальных ядер, гипоталамуса и ствола мозга. Эти волокна обозначены на рисунке ниже зеленым цветом. Несмотря на то, что они распространены по всей коре, их возбуждение не сопровождается генерализованной или неспецифической реакцией. Анатомическая специфичность волокон (корковая, пластинчатая и клеточная) обусловливает возбуждение или торможение только определенных групп нейронов. Ядра их начала и нейромедиаторы: д) Эфферентные волокна. Единственные эфферентные волокна, покидающие кору больших полушарий, представлены возбуждающими аксонами пирамидных клеток. Одна часть аксонов пирамидных клеток соединяется с длинными и короткими ассоциативными волокнами, другая образует комиссуральные или проекционные пути. Ассоциативные и комиссуральные пути составляют большой объем белого вещества полушарий головного мозга. • Примерами коротких ассоциативных путей (проходят между соседними зонами коры в составе поверхностного белого вещества в виде U-волокон) служат пути, направляющиеся в моторную кору от сенсорной коры и обратно. Примерами длинных ассоциативных путей служат волокна между префронтальной корой (лежащей кпереди от моторных зон коры) и ассоциативными сенсорными зонами. Источник этих волокон — пирамидные клетки, расположенные преимущественно в слоях II и III. • Комиссуральные волокна мозга представлены исключительно аксонами пирамидных клеток, проходящих через мозолистое тело, переднюю и заднюю спайки (и другие мелкие спайки) к аналогичным участкам противоположного полушария (например, волокна от первичной корковой зоны направляются к соответствующей ассоциативной области другого полушария), а также к несоответствующим зонам [подобные комиссуральные связи отсутствуют между первичной зрительной корой, первичной соматосенсорной и моторной корой, иннервирующими дистальный отдел верхней конечности]. Волокна начинаются от пирамидных клеток, расположенных преимущественно в слоях II и III. • Основная часть проекционных волокон от первичной сенсорной и моторной коры подходит к базальным ганглиям. К таламусу направляются волокна от всех отделов коры. К другим основным проводящим путям относят корково-мостовой (к ипсилатеральным ядрам моста), корково-ядерный (к двигательным и чувствительным ядрам черепных нервов моста и продолговатого мозга на противоположной стороне) и корково-спинномозговой. Источником этих волокон служат пирамидные клетки, расположенные преимущественно в слоях V и VI (отдающие волокна преимущественно к специфическим релейным ядрам таламуса). — Также рекомендуем «Корковые зоны головного мозга: цитоархитектонические поля Бродмана, методы исследования» Редактор: Искандер Милевски. Дата публикации: 22.11.2018 |
Источник
