Судьба молочной кислоты цикл кори
В клетках организма всегда существует потребность в глюкозе:
- для эритроцитов глюкоза является единственным источником энергии,
- нервная ткань потребляет около 120 г глюкозы в сутки и эта величина практически не зависит от интенсивности ее работы. Только в экстремальных ситуациях (длительное голодание) она способна получать энергию из неуглеводных источников (кетоновые тела),
- глюкоза играет весомую роль для поддержания необходимых концентраций метаболитов цикла трикарбоновых кислот (в первую очередь оксалоацетата).
Таким образом, при определенных ситуациях – при низком содержании углеводов в пище, голодании, длительной физической работе, т.е. когда глюкоза крови расходуется и наступает гипогликемия, организм должен иметь возможность синтезировать глюкозу и нормализовать ее концентрацию в крови. Это достигается реакциями глюконеогенеза, идущими в печени.
По определению, глюконеогенез – это синтез глюкозы из неуглеводных компонентов: лактата, пирувата, глицерола, кетокислот цикла Кребса и других кетокислот, из аминокислот.
Необходимость глюконеогенеза и его значение для организма демонстрируют два цикла – глюкозо-лактатный и глюкозо-аланиновый.
Глюкозо-лактатный цикл (цикл Кори)
Глюкозо-лактатный цикл – это циклический процесс, объединяющий реакции глюконеогенеза и реакции анаэробного гликолиза. Глюконеогенез происходит в печени, субстратом для синтеза глюкозы является лактат, поступающий в основном из эритроцитов или мышечной ткани.
В эритроцитах молочная кислота образуется непрерывно, так как для них анаэробный гликолиз является единственным способом образования энергии.
В скелетных мышцах высокое накопление молочной кислоты (лактата) является следствием гликолиза при очень интенсивной, субмаксимальной мощности, работе, при этом внутриклеточный рН снижается до 6,3-6,5. Но даже при работе низкой и средней интенсивности в скелетной мышце всегда образуется некоторое количество лактата.
Убрать молочную кислоту можно только одним способом – превратить ее в пировиноградную кислоту. Однако сама мышечная клетка ни при работе, ни во время отдыха не способна превратить лактат в пируват из-за особенностей изофермента лактатдегидрогеназы-5. Зато клеточная мембрана высоко проницаема для лактата и он движется по градиенту концентрации наружу. Поэтому во время и после нагрузки (при восстановлении) лактат легко удаляется из мышцы. Это происходит довольно быстро, всего через 0,5-1,5 часа в мышце лактата уже нет. Малая часть молочной кислоты выводится с мочой.
Большая часть лактата крови захватывается гепатоцитами, окисляется в пировиноградную кислоту и вступает на путь глюконеогенеза. Глюкоза, образованная в печени, используется самим гепатоцитом или возвращается обратно в мышцы, восстанавливая во время отдыха запасы гликогена. Также она может распределиться по другим органам.
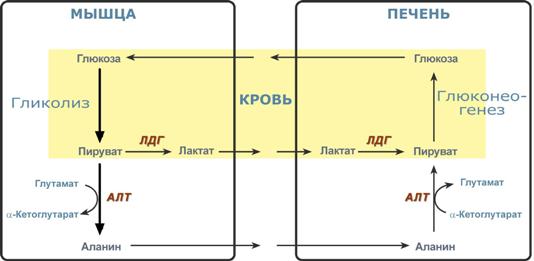
Глюкозо-лактатный (выделен желтым) и глюкозо-аланиновый циклы
Глюкозо-аланиновый цикл
Целью глюкозо-аланинового цикла также является уборка пирувата, но кроме этого решается еще одна немаловажная задача – доставкааминного азота из мышцы в печень.
При мышечной работе и в покое в миоците распадаются белки и образуемые аминокислоты трансаминируются с α-кетоглутаратом и полученный глутамат взаимодействует с пируватом. Образующийся аланин является транспортной формой аминного (аминокислотного) азота и пирувата из мышцы в печень. В гепатоците идет обратная реакция трансаминирования, аминогруппа через глутамат передается на синтез мочевины, пируват используется для синтеза глюкозы.
Кроме мышечной работы, глюкозо-аланиновый цикл активируется во время голодания, когда белки мышц и других тканей распадаются и многие аминокислоты используются в качестве источника энергии, а их азот необходимо доставить в печень.
Источник
Лактат – конечный продукт анаэробного окисления глюкозы в мышцах, особенно в белых мышечных волокнах, где митохондрий меньше, чем в красных. Может включаться в глюконеогенез после окисления до пирувата в лактатдегидрогеназной реакции. При продолжительной физической работе основным источником лактата является скелетная мускулатура, в клетках которой преобладают анаэробные процессы. Накопление молочной кислоты в мышцах ограничивает их работоспособность. Это связано с тем, что при повышении концентрации молочной кислоты в ткани снижается уровень рН (молочнокислый ацидоз). Изменение рН приводит к ингибированию ферментов важнейших метаболических путей. В утилизации образующейся молочной кислоты важное место принадлежитглюкозо-лактатному циклу Кори.
Цикл Кори и глюкозо-аланиновый цикл (пояснения в тексте).
Лактат, образовавшийся в мышцах, переносится кровью в печень, где в процессе глюконеогенеза превращается в глюкозу, которая с током крови может возвращаться в работающую мышцу. В печени часть лактата может окисляться до углекислого газа и воды, превращаться в пируват и вовлекаться в общий путь катаболизма.
Значение цикла Кори:
1. Регуляция постоянного уровня глюкозы в крови.
2. Обеспечивает утилизацию лактата.
3. Предотвращает накопление лактата (снижение рН — лактоацидоз).
4. Экономичное использование углеводов организмом.
Регуляция обмена углеводов осуществляется на уровне тканей – кровь, печень, мышцы.
Глюкогенные аминокислоты, к которым относятся большинство белковых аминокислот. Ведущее место в глюконеогенезе среди аминокислот принадлежит аланину, который может превращаться в пируват путём трансаминирования. При голодании, физической работе и других состояниях в организме функционирует глюкозо-аланиновый цикл, подобный циклу Кори для лактата (рисунок 16.2). Существование цикла аланин – глюкоза препятствует отравлению организма, так как в мышцах нет ферментов, утилизирующих аммиак. В результате тренировки мощность этого цикла значительно возрастает.
Другие аминокислоты могут, подобно аланину, превращаться в пируват, а также в промежуточные продукты цикла Кребса (α-кетоглутарат, фумарат, сукцинил-КоА). Все эти метаболиты способны преобразовываться в оксалоацетат и включаться в глюконеогенез.
Глицерол – продукт гидролиза липидов в жировой ткани.Этот процесс значительно усиливается при голодании. В печени глицерол превращается в диоксиацетонфосфат – промежуточный продукт гликолиза и может быть использован в глюконеогенезе.
Жирные кислоты и ацетил-КоА не являются предшественниками глюкозы. Окисление этих соединений обеспечивает энергией процесс синтеза глюкозы.
Энергетический баланс. Путь синтеза глюкозы из пирувата (рисунок 16.6) содержит три реакции, сопровождающиеся потреблением энергии АТФ или ГТФ:
а) образование оксалоацетата из пирувата (затрачивается молекула АТФ);
б) образование фосфоенолпирувата из оксалоацетата (затрачивается молекула ГТФ);
в) обращение первого субстратного фосфорилирования – образование 1,3-дифосфоглицерата из 3-фосфоглицерата (затрачивается молекула АТФ).
Каждая из этих реакций повторяется дважды, так как для образования 1 молекулы глюкозы (С6) используются 2 молекулы пирувата (С3). Поэтому энергетический баланс синтеза глюкозы из пирувата составляет – 6 молекул нуклеозидтрифосфатов (4 молекулы АТФ и 2 молекулы ГТФ). При использовании других предшественников энергетический баланс биосинтеза глюкозы отличается.
Источник
ОБМЕН УГЛЕВОДОВ
Классификация:
1) Моносахариды – глюкоза, фруктоза, галактоза.
2) Олигосахариды – мальтоза, сахароза, лактоза.
3) Полисахариды – крахмал и гликоген.
Гликоген – полисахарид в котором молекулы глюкозы соединены 1,4-гликозидными связями, а в местах ветвления 1,6-гликозидными связями.
Крахмал – гетерополисахарид, состоящий из линейной амилозы и разветвленного амилопектина. Строительная единица крахмала – глюкоза соединенная 1,4- и 1,6-гликозидными связями.
Переваривание углеводов:
Начинается в ротовой полости, под действием амилазы слюны, которая является эндоамилазой, то есть разрывает внутренные 1,4-гликозидные связи. Действие про pH=6,8-7.0
Амилаза в активном центре содержит Ca, активатором амилазы является анион Cl-.
Глубокого переваривания в ротовой полости не происходит, так как пища там находится недолго. Образуются промежуточные продукты гидролиза крахмала – декстрины(амило-, эритро-, мальто-, ахродекстрины).
При попадании пищевого комка в желудок амилаза слюны инактивируется, так как pH желудочного сока 1,5-2,5
Переваривание углеводов внутри пищевого комка не происходит, так как соляная кислота не попадает внутрь пищевого комка. Основное переваривание углеводов происходит в кишке под действием панкреатической амилазы(диастаза), которая разрывает внутренние 1,4-гликозидные связи.
Панкреатический сок содержит еще 2 фермента – Амило- и олиго-1,6-гликозидазы, которые разрывают 1,6-гликозидные связи.Конечными продуктами переваривания крахмала являются мальтоза, мальтотриоза, глюкоза.
Кишечный сок содержит сахаразу, которая действует на сахарозу и расщепляет её на сахарозу и фруктозу. Так же в нем есть мальтаза, которая действует на мальтозу, расщепляя ее на 2 остатка глюкозы. Лактаза действует на лактозу и расщепляет ее на глюкозу и галактозу.
Конечные продукты переваривания полисахаридов являются моносахариды. Образовавшиеся моносахариды всасываются с различной скоростью, скорость всасывания глюкозы 100 процентов.
Использование глюкозы .
1) синтез гликогена в печени и скелетных мышцах. Наибольшее количество гликогена откладывается в мышцах, запас гликогена хватает на 24 часа. Гликоген откладывается в виде гранул или глыбок, где содержаться ферменты и синтеза и распада гликогена.
Глюкоза, попав в клетку активируется или фосфорилируется с образованием глюкозо-6-фосфата. Эту реакции катализирует ферменты – глюкокиназа или гексокиназа, которая обладает различным сродством к глюкозе. Реакция не обратимая и требует АТФ. Глюкозо-6-фосфат основной метаболит обмена углеводов.
2) Глюкозо-6-фосфат(фермент изомераза)→ глюкоза-1-фосфат.
3) Глюкоза-1-фосфат взаимодействует с УТФ в результате образуется транспортная форма глюкозы при синтезе гликогена УДФ-глюкоза. Фермент реакции трансфераза.
4) УДФ-глюкоза передает свой остаток глюкоза-1-фосфат на затравочное количество гликогена с образование 1,4-гликозидной связи. Фермент гликогенсинтетаза – главный фермент синтеза гликогена, может существовать в двух формах: фосфорилированная неактивная, дефосфорилированная активная. В образовании 1,6-гликозидных связей участвует фермент ветвления.
Распад Гликогена
Протекает при участии фермента гликоген-фосфарилазы, который может находится в 2 формах:
-фосфорилированная активная
-дефосфорилированная неактивная
Активации гликоген-фосфорилазы осуществляют гормоны адреналин и глюкагон.
Адреналин действует при стрессах , активирует распад гликогенав печени и скелетных мышцах. Глюкагон действует в норме и в постабсортивный период, активируя распад гликогена только в печени.
Механизм действия глюкагона и адреналина
Так как для этих гормонов мембрана клетки не проницаема, рецепторы для них располагается на внешней поверхности клеточной мембраны. Происходит образование гормон-рецепторного комплекса, затем изменяется конформация G-белка за счет гидролиза ГТФ в ГДФ. G-белок расположен в самой мембране.
Активируется аденилатциклаза, расположенная на внутренней поверхности мембраны. Активированная аденилатциклаза в клетке катализирует реакцию образования цАМФ из АТФ.
цАМФ вторичный посредник или мессенджер в передаче гормонального сигнала внутрь клетки. На этом этапе происходит усиление гормонального сигнала цАМФ образуется много до 500 молекул, но время их жизни очень мало! так как они быстро гидролизируются ферментами. цАМФ активирует протеинкиназу, актиная протеинкиназа фосфорилирует белки-ферменты клетки. Активная гликоген-фосфорилаза отщепляет от гликогена глюкоза-1-фосфат→глюкозо-6-фосфат→глюкоза→в кровь.
Гликолиз
Протекает в цитоплазме может быть аэробный и анаэробный.
Анаэробный гликолиз – это окисление глюкозы в отсутствии кислорода до двух молекул лактата и 2-х АТФ.
1) Образование глюкозо-6-фосфат. Реакция необратима, затрачивается 1 молекула АТФ.
2) Глюкозо-6-фосфат(мутаза)→фруктозо-6-фосфат
3) Фруктозо-6-фосфат при участии АТФ и фермента фосфофруктокиназы фосфорилируется с образованием фруктоза-1,6—дифосфат. Реакция необратима, затрачивается АТФ. Эта реакция определяет скорость гликолиза в целом. Фосфофруктокиназа алостерический фермент его активность угнетается высоким содержанием АТФ и повышается при высоких концентрациях АМФ в клетке.
4) Фруктоза-1,6-фосфат при участии фермента альдолаза распадается на 2 фосфотриозы: диоксиацетонфосфат, глицероальдегидтрифосфат.
5)Диоксиацетонфосфат(триозофосфоизомераза)→ глицероальдегидтрифосфат.
На этом заканчивается неокислительная стадия гликолиза и начинается стадия гликолитической оксиредукции, в которую вступают 2 молекулы глицероальдегиттрифосфат.
6) Реакция окислительного фосфорилирования протекает при участии неорганического фосфата. (H3PO4) НАД и фермента глицероальдегидфосфатдегидрогиназа. В результате образуется 1,3-дифосфоглицерат + 2НАДН2.
7) Субстратное фосфорилирование. 1,3-дифосфоглицерат передает свой остаток фосфата из первого положения на АДФ в результате образуется 2 молекулы АТФ и 2 молекул 3-фосфоглицерата. Фермент фосфоглицераткиназа.
8) 3-фосфоглицерат(мутаза)→2-фосфоглицерат.
9) 2-фосфоглицерат(енолаза)→фосфоенолпируват содержит макроэргическую связь, обозначаемую значком ~.
10) Субстратное фосфорилирование. 2-фосфоенолпируват + 2АДФ→2пируват + 2АТФ. Фермент пируваткиназа, реакция необратима.
11) При участии НАДН2, которые образовались в 6-й реакции, восстанавливается в лактат. Фермент лактат дегидрогиназа.
Энергетический выход анаэробного гликолиза:
1-3-й реакции затрачивается 2 молекулы АТФ. 7,10-й реакции образуется 4 молекулы АТФ. И того 4-2=2 молекулы АТФ.
Биологическое значение анаэробного гликолиза.
Заключается в том что клетка получает 2 молекулы АТФ в отсутствии кислорода.
Аэробный гликолиз – это окисление глюкозы в присутствии кислорода с образованием 2 молекул пирувата и 7 молекул АТФ. Аэробный гликолиз состоит из 10 реакций. Энергетический выход аэробного гликолиза:
1-3-й реакции затрачивается 2 молекулы АТФ. 6-й реакции 2 молекулы НАДН2, которые поступают в дыхательную цепь и там дают 2.5 молекулы АТФ*2=5 молекул АТФ.(одна молекула НАДН2 дает 2.5 молекул АТФ, а одна молекула ФАДН2 дает 1.5 молекулы АТФ). 7,10-й реакции образуется 4 молекулы АТФ. Итого 5+4-2=7 молекул АТФ.
Анаэробный гликолиз отличается от аэробного гликолиза:
1) условие проведения
2) количество реакций анаэробный – 11, аэробный – 10.
3) Конечные продукты анаэробный – 2 молекулы лактата, аэробный – 2 молекулы пирувата.
4) Количество энергии анаэробный – 2 молекулы АТФ, аэробный – 7 молекул АТФ.
Глюконеогенез
Это синтез глюкозы из неуглеводных продуктов – пируват, лактат, гликогенные аминокислоты(белки), глицерин(жиры). Синтезировать глюкозу из пирувата и лактата по пути гликолиза невозможно, так как 3 реакции гликолиза являются необратимыми 1,3,10. Поэтому в глюконеогенезе обходные пути и специальные ферменты. Например, надо синтезировать глюкозу из 2-х молекул пирувата. Пируват не может превратится в фосфоенолпируват по пути гликолиза, так как 10-я реакция необратима.
1-я обходная реакция пируват в митохондриях при участии 2-х молекул АТФ и CO2 карбоксилируется в оксалоацетат. Фермент этой реакции пируваткарбоксилаза, Кофермент витамин Н(биотин), при нехватки витамина активность фермента снижается. Оксалоацетат не может попасть в цитозоль, так как мембрана митохондрий для него не проницаема, сначала оксалоацетат превращается в малат, который переходит в цитозоль и вновь превращается в оксалоацетат. Оксалоацетат при участии 2-х молекул ГТФ и фермента фосфоенолпируваткарбоксикиназа превращается в фосфоенолпируват. Далее идут реакции гликолиза фосфоенолпируват→2-фосфоглицерат→3-фосфоглицерат(2АТФ) →1,3-дифосфоглицерат→2 молекулы глицероальдегидтрифосфат (1 молекула дает фрутозо-1,6-дифосфат) другая молекула дает диоксиацетонфосфат→фрутозо-1,6-дифосфат, который не может превратится во фрутоза-6-фосфат по пути гликолиза, так как 3-я реакция гликолиза необратима.
2-я обходная реакция ее катализирует фермент фруктозодифосфотаза при участии которого фрутозо-1,6-дифосфат превращается во фруктозо-6-фосфат→глюкозо-6-фосфат, который не может превратится в свободную глюкозу по пути гликолиза, так как 1-я реакция гликолиза необратима.
3-я обходная реакция катализирует фермент глюкозо-6-фосфатаза. На синтез 1 молекулы глюкозы из 2-х молекул пирувата тратится 6 молекул АТФ. Глюконеогенез контролтрует кортизол.
Глюкозолактатный цикл(цикл Кори).
Гликолиз, который протекает в скелетных мышцах связан с глюконеогенезом, который протекает в печени. При мышечной работе в них накапливается лактат, который выходит в кровь и попадает в печень, где из него синтезируется глюкоза путем глюконеогенеза. Эта глюкоза выходит в кровь, затем попадает в мышцы где используется в качестве энергетического материала.
Пировиноградная кислота, которая образовалась в результате аэробного гликолиза, подвергается окислительному карбоксилированию с образованием ацетилКоА и НАДН2(2.5 АТФ). Эту реакцию катализирует пируватдегидрогиназный комплекс, который состоит из 3 ферментов и 5 коферментов.
3 фермента:
пируватдекарбоксилаза,липоацетилтрансфераза, липоамиддегидрогиназа.
5 коферментов: Тиаминпирофосфат, который связан с пируватдекарбоксилазой , липоат(липоевая кислота), Коэнзим А, ФАД, НАД.
Источник
Дано определение молочной кислоты, описана история ее открытия и метаболизм ее превращения в организме при физических нагрузках (цикл Кори). Описывается концепция ацидоза, описывающая изменения в скелетных мышцах, которые приводят к их гипертрофии и увеличению силовых показателей.
Молочная кислота (лактат) и физические нагрузки
Определение
Молочная кислота (лактат) – конечный продукт анаэробного распада глюкозы и гликогена (гликолиза).
История открытия
1780 году шведский химик Карл Вильгельм Шилле выделил молочную кислоту из молока. А в 1808 году Йенс Якоб Берцелиус открыл, что молочная кислота образуется в скелетных мышцах при выполнении физических упражнений.
Молочная кислота и физические нагрузки
Практически при любой физической нагрузке для получения АТФ используется гликоген скелетных мышц. Его концентрация в скелетных мышцах при интенсивных физических нагрузках быстро снижается. Одновременно в скелетных мышцах образуется и накапливается молочная кислота.
Формула молочной кислоты (С3H6O3).
Цикл Кори
Циклический путь метаболизма молочной кислоты в скелетных мышцах открыт американским биохимиком, нобелевским лауреатом Герти Терезой Кори. По другим источникам открытие цикла Кори приписывается нобелевским лауреатом супругам Карлу и Герти Кори. Он описывает превращения молочной кислоты в организме человека. Большая часть молочной кислоты, которая образуется в организме во время физических нагрузок включается в метаболические процессы непосредственно в мышцах и под влиянием фермента лактатдегидрогеназы (ЛДГ) превращается в пировиноградную кислоту, которая затем в митохондриях окисляется до углекислого газа и воды. Другая часть молочной кислоты через кровеносные капилляры проникает в кровь и доставляется в печень, где включается в метаболические реакции, приводящие к синтезу гликогена. Незначительное количество молочной кислоты может выводиться из организма с мочой и потом. Гликоген печени используется организмом для восстановления энергетических источников скелетных мышц.
Концепция ацидоза
Одной из концепций, объясняющей возникновение острых болезненных ощущений, возникающих как во время, так и после тренировки, является предположение, что накопление молочной кислоты в мышечных волокнах является пусковым механизмом целого ряда биохимических реакций.
Во-первых, молочная кислота повышает кислотность внутри мышечных волокон. Изменение рН саркоплазмы мышечных волокон с 7,1 до 6,5 (то есть повышение кислотности) при сильном утомлении снижает активность ключевых ферментов гликолиза – фосфорилазы и фосфофруктокиназы. При значении рН саркоплазмы равном 6,4 расщепление гликогена прекращается. Это вызывает резкое снижение уровня АТФ и развитие утомления (Н.И. Волков с соавт., 2000).
Во-вторых, повышение концентрации молочной кислоты в мышечных волокнах приводит к увеличению проницаемости их мембраны, повышению осмотического давления, в результате чего в мышечные волокна поступает вода. Возникает отёк, мышечные волокна «разбухают» и сдавливают болевые рецепторы мышц. Это ощущается как «жжение» в мышцах (М.И. Калинский, В.А. Рогозкин, 1989). Спортсмены называют это явление «мышцы забиты».
Молочная кислота и лактат
Следует отметить, что молочная кислота и лактат — не одно и то же. Лактат — это соль молочной кислоты. Образовавшаяся в результате гликолиза в скелетных мышцах молочная кислота почти полностью диссоциирует на ионы водорода и соединение, которое соединяется с ионами натрия или калия и образует соль (лактат), рис. 1.
Рис. 1
Поэтому в литературе часто вместо понятия «молочная кислота» используется термин «лактат». Содержание молочной кислоты и лактата имеет взаимосвязь с кислотностью внутри мышечных волокон (то есть с pH саркоплазмы). При pH в интервале от 6.5 (полное утомление) до 7,1 (норма) в мышечных волокнах накапливается, выводится и перерабатывается именно лактат.
В течение нескольких часов лактат удаляется из мышечных волокон. Если после физической нагрузки выполнить 10-15 минутную аэробную работу (например, бег или педалирование на велосипеде), лактат из мышц выведется еще быстрее.
Новые исследования свидетельствуют о том, что основным повреждающим агентом являются ионы водорода (Н+). Утомление в скелетных мышцах вызывает ацидоз — накопление ионов водорода и смещение pH саркоплазмы в кислую сторону (R. A. Robergs et al., 2004).
Молочная кислота, гипертрофия и сила скелетных мышц
Предполагается, что накопление кислых продуктов в мышечных волокнах (ацидоз) лежит в основе их повреждения, что в последствии приводит к их гипертрофии по миофибриллярному типу и росту силы. Следовательно, удалять молочную кислоту из скелетных мышц после тренировки не следует, так как это основной фактор, повреждающий мышечные волокна. Это предположение подтверждается опытом тренировок чемпионки мира в беге на 400 м с барьерами Марины Степановой и ее тренера Вячеслава Владимировича Степанова. Стремясь увеличить силовые показатели мышц ног, М. Степанова и В. Степанов в цикле своих статей «Анаэробика» указывают, что «есть смысл ненадолго (на несколько часов) «повариться» в молочнокислой среде, а «разогнать» ее позже (к примеру, вечерними упражнениями)».
Литература
- Калинский, М.И. Биохимия мышечной деятельности / М.И. Калинский, В.А. Рогозкин. – Киев: Здоровья, 1989.– 144 с.
- Михайлов С.С. Спортивная биохимия. – М.: Советский спорт, 2009.– 348 с.
- Самсонова, А.В. Гипертрофия скелетных мышц человека: Учеб. пособие. 5-е изд. /А.В. Самсонова. – СПб: Кинетика, 2018.– 159 с.
- Самсонова, А. В. Гормоны и гипертрофия скелетных мышц человека: Учеб. пособие. – СПб: Кинетика, 2019.– 204 c.: ил.
- Степанова, М. Анаэробика /М. Степанова, В. Степанов // Легкая атлетика, 2011 № 7-8. С. 24-27.
С уважением, А.В. Самсонова
Источник

